
Hippocampus
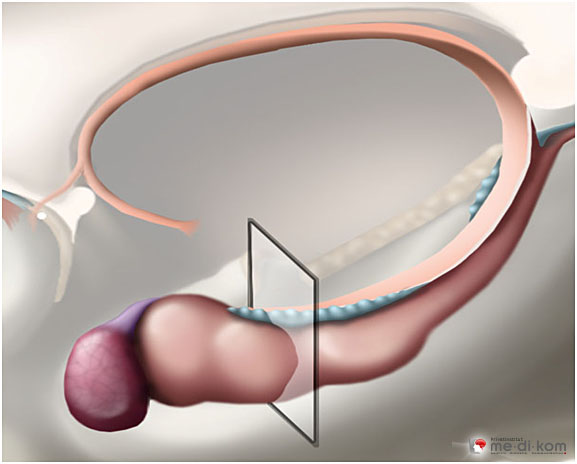
Der Hippocampus liegt halbmondförmig gekrümmt am inneren Rand des Schläfenlappens. Er spielt eine große Rolle für Lernen und Gedächtnis, insbesondere bei der Überführung von Lerninhalten vom Kurzzeit- ins Langzeitgedächtnis, d. h. der langfristigen Gedächtnisbildung. Hierfür vergleicht er neu ankommende und relevante mit bereits gespeicherten Informationen. Der Hippocampus gliedert sich in drei Bereiche:
- Ammonshorn (Cornu ammonis) mit den Feldern CA1-CA4.
- Gyrus dentatus
- Subiculum
Unter Einbeziehung folgender weiterer Strukturen der Umgebung spricht man auch von der Hippocampusformation:
- Area entorhinalis
- Area perirhinalis
- Praesubiculum
- Area parahippocampalis caudalis
Das Ammonshorn besteht aus den Subregionen CA1-CA4. Seine großen Pyramidenzellen erhalten Afferenzen v.a. aus dem entorhinalen Kortex (über den Gyrus dentatus). Darüber hinaus erhalten diese Zellen modulierende Eingänge aus den Raphekernen, dem Locus caeruleus, Nucleus incertus und Septum.
Der Gyrus dentatus gilt aufgrund seiner zahlreichen kortikalen Afferenzen aus dem entorhinalen Kortex als wichtige Eingangsstruktur.
Das Subiculum stellt die wichtigste Ausgangsstation des Hippocampus dar. Die Axone seiner Pyramidenzellen und der CA3-Region bilden mit Kollateralen Verbindungen, die einerseits über den Fornix zum Septum ziehen, andererseits zum entorhinalen Kortex führen, der wieder mit dem Isokortex verschaltet ist.
Der entorhinale Kortex gilt als bedeutsamster Eingang der Hippocampusformation, denn ihn erreichen Afferenzen aus zahlreichen (vielfach assoziativen) Arealen.
Der perirhinale Kortex vermittelt Signale zwischen Hippocampus und Isokortex.
Das Praesubiculum markiert die Grenze zwischen entorhinalem Kortex und Subiculum.
Insgesamt kann der Hippocampus als wichtiger Knotenpunkt hier einstrahlender neo- und subkortikaler Fasern angesehen werden. Es treffen vier reziproke Schaltkreise zusammen (Roth und Dicke, 2005):
- Zwischen entorhinalem Kortex (der wiederum Zuflüsse aus vielen neokortikalen Gebieten erhält) und Hippocampus
- Zwischen Amygdala ('emotionales' Gedächtnis) und Hippocampus
- Zwischen Mammillarkörper und Hippocampus (Papez-Kreis)
- Zwischen septaler und präoptisch-hypothalamischer Region und Hippocampus (wobei das Septum als Verlängerung des aufsteigenden Aktivierungssysems des Hirnstamms angesehen werden kann)
Hier werden die externen (sensorischen) Informationen mit internen Informationen abgeglichen und verknüpft. Bahnen aus den primären visuellen, akustischen und somatosensiblen Arealen des Neokortex, die über zwischengeschaltete Assoziationsgebiete zur hippocampalen Formation ziehen, versorgen den Hippocampus mit sensorischer Information aller Modalitäten. Der Großteil dieser Fasern endet in der Area entorhinalis. D.h. Afferenzen des Isokortex erreichen den Hippocampus nach Umschaltung im entorhinalen Kortex. Innerhalb des Hippocampus zeigt sich schließlich ein charakteristisches Verschaltungsmuster.
Eine Bildserie stellt dieses anschaulich dar, indem die Verschaltungsprinzipien schrittweise aufgebaut werden.
|
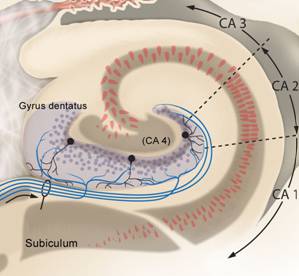
Das Querschnittsbild zeigt die unterschiedlichen Felder innerhalb der Hippocampus-Formation: Gyrus dentatus, CA4, CA3, CA2, CA1 und Subiculum. |
|
Die Afferenzen aus zahlreichen kortikalen Arealen kommen über den Entorhinalen Kortex (EC) in Form des Tractus perforans an den Körnerzellen des Gyrus dentatus an. |
|
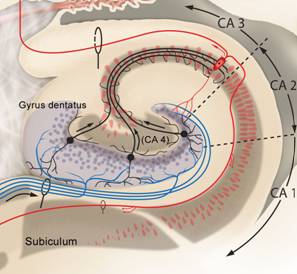
Von den Körnerzellen des Gyrus dentatus ziehen die Moosfasern zu Zellen der CA3-Region. |

|
Die Zellen der CA3-Region projizieren mit ihren Schaffer-Kollateralen zu Zellen der CA1-Region. |
|
Die CA1-Zellen senden ihre Fasern u.a. über das Subiculum zurück zum Kortex. Der Schaltkreis Kortex-EC-Hippocampus -Kortex wird geschlossen. |
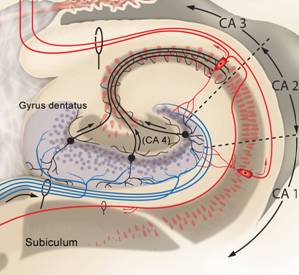
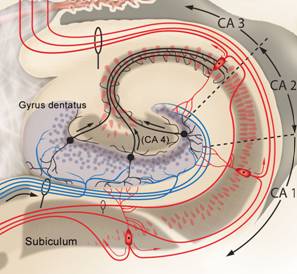
Nach den Bildern nun eine Zusammenfassung in Worten: Die glutamatergen Zuflüsse verlaufen im Tractus perforans und münden an den Körnerzellendendriten des Gyrus dentatus. Von dort verlaufen Axone der gleichfalls mit Glutamat arbeitenden Körnerzellen als Moosfasern zur Region CA3, und zwar zu den Dendriten der dortigen Pyramidenzellen. Diese ebenso glutamatergen Neurone sind über die Schaffer-Kollateralen mit Dendriten der Pyramidenzellen in Region CA1 verbunden. Von dort führen Axone über das Subiculum wieder zum entorhinalen Kortex. Nach einer intrahippocampalen tri-synaptischen Verschaltung erfolgt die Rückprojektion zum entorhinalen Kortex. Somit ist der Kreislauf entorhinaler Kortex - Hippocampus - entorhinaler Kortex geschlossen und die verarbeiteten Informationen werden als Reafferenzkopie wieder zum Isokortex zurückgesandt. So dass der Kreislauf eigentlich insgesamt als Kortex-EC-Hippocampus-EC-Kortex – Schleife beschrieben werden kann (Zilles, 2005, S. 121).
Die beschriebenen glutamatergen Verschaltungen bilden ein System, das für Lernfunktionen zentral ist. Denn hier hat man im Zusammenhang mit den glutamatergen NMDA-Rezeptoren Langzeit-potenzierungen festgestellt, d. h. anhaltende Veränderungen der synaptischen Übertragungsmuster. Sie stellen eine interne Repräsentation von Gedächtnisinhalten dar und können als neuronales Korrelat der Gedächtniskonsolidierung angesehen werden (ebd., S. 122).
Ferner finden sich in der hippocampalen Formation mehrere Klassen inhibitorischer Neurone, die GABA als Neurotransmitter nutzen und einen starken Effekt auf die Netzwerkfunktion (Rhythmus der Erregungsfrequnz) des Hippocampus ausüben.
Daneben erhält der Hippocampus zahlreiche modulierende Eingänge aus dem Septum (cholinerg), den Raphe-Kernen (serotonerg), dem Mamillarkörper (histaminerg) und dem Locus caeruleus (noradrenerg).
Neurovulnerabilität des Hippocampus bei Psychotrauma
In mehrfach replizierten Studien mit Bildgebungsverfahren (Kernspinresonanztomo-graphie) wurden wiederholt um 5-26 Prozent verkleinerte Hippocampusformationen bei PTBS-Patienten im Vergleich zu gesunden Probanden festgestellt (Bremner, 1995, Bremner et al., 1997; Gurvits et al., 1996; Stein et al. 1997 zit. in Bering, 2005). Diese Veränderungen traten jedoch uneinheitlich ein- oder beidseitig auf und sind auch insgesamt nicht ohne widersprüchliche Befunde geblieben (vgl. Bering, 2005, S. 121 ff.).
Allerdings ist die Hippocampusatrophie auch für andere Erkrankungen wie Schizophrenie und Depressionen nachgewiesen. Hierbei ist zu berücksichtigen, dass z. B. depressive u. a. Störungen, die eine Atrophie begünstigen, als komorbide Störungen von Psychotraumatisierungen auftreten.
Neuere und sensitivere Messmethoden weisen aber auf feinstrukturelle Veränderungen in der Hippocampusregion hin, selbst wenn noch keine Volumenänderung nachweisbar ist. Die Kernspinprotonenspektroskopie erlaubt es, die Menge eines Moleküls in einer bestimmten Hirnregion zu erkennen. Zum Beispiel fungiert NAA (N-Acetyl-Aspartat) als neuronaler Integritätsmarker: Studien ergaben bei PTBS-Patienten durchschnittlich 23 Prozent weniger NAA. Dies ist als Hinweis darauf zu werten, dass in ihrem Fall bereits neuronaler Zellverlust vorliegt (Roth, 2007).
Um der Frage nachzugehen, ob es sich bei diesen Veränderungen der Hippocampus-formation um Ursache (Hypothese [H1]: Volumenminderung als hereditärer Faktor), Begleiterscheinung (Hypothese [H2]: Volumenminderung ausgelöst durch traumatische Situation) oder Folge einer psychischen Traumatisierung handelt (Hypothese [H3]: Neurovulnerabilität durch traumatischen Prozess verursacht), schlägt Bering (2005) vor, die unterschiedlichen Befunde und Hypothesen zur hippocampalen Neurovulnerabilität auf Grundlage des Verlaufsmodell zu betrachten. Abgestimmt auf die Phasen des Verlaufsmodells psychischer Traumatisierung geht er von einer multikausalen Entstehung aus und sucht die verschiedenen Hypothesen zu integrieren (vgl. Bering, 2005, 141 ff.).
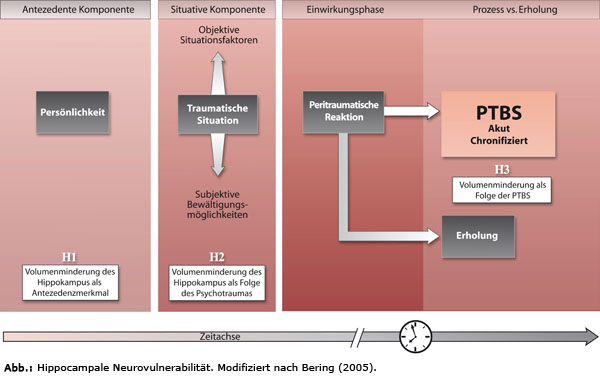
Dabei verknüpft Bering die Ergebnisse aus der experimentellen Wirkungsforschung mit der prozessorientierten Verlaufsbetrachtung (ebd., S. 365). Die „Dysregulation der Stressachse“ (ebd.) stellt eines der analysierten Konzepte dar.
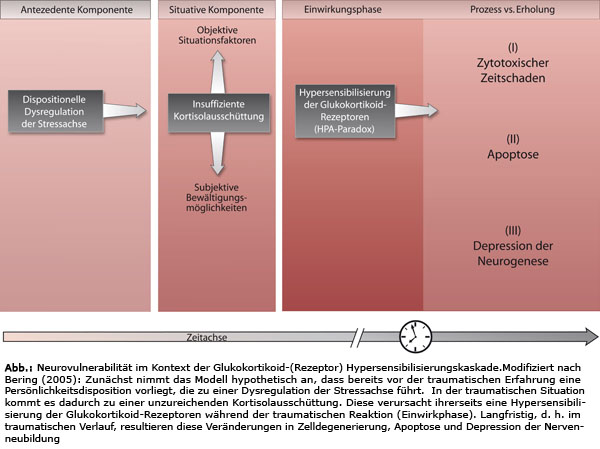
Die Schlussfolgerungen von Bering (2005, S. 365) werden hier im Original wiedergegben:
- „Die Ursachenforschung zur Erklärung der Hypovolämie des Hippocampus bei Psychotraumastörungen sollte nicht auf eine Momentaufnahme der hereditären, situativen oder Prozessfaktoren fokussieren, sondern auf Denkmodelle eines interdisziplinären komplex-kybernetischen Verlaufsprozesses zurückgreifen.
- Argumente verdichten sich, dass die Hypovolämie der Hippokampusformation auch bei der PTBS auf einen Pathomechanismus zurückzuführen ist, der – im Sinne der Pathoklise – an charakteristische physikochemische Eigenschaften bestimmter Zellpopulationen in der Hippokampusformation gebunden ist.
- Die Ergebnisse zeigen, dass die PTBS nicht mit dem Hyperkortisolmodell der Stressforschung vereinbar ist. Die Hypersensibilisierung des Glukokortikoidrezeptors könnte eine Erklärung sein, warum ein niedriges Kortisol bei PTBS- Patienten die Vulnerabilität hippokampaler Neuronenverbände steigert.
- Für die Vulnerabilität von Neuronen bei Psychotraumastörungen müssen pathophysiologische Mechanismen des zytotoxischen Zellschadens, der Apoptose und der verminderten Neurogenese der Körnerzellen in Betracht gezogen werden“ (ebd.).


